Botrytis cinerea, commonly known as grey mould, is a necrotrophic polyphagous pathogen belonging to the phylum Ascomycota. B. cinerea has a remarkable ability to infect a wide range of plant species, similar to Sclerotinia sclerotiorum, which causes white mould disease in diverse plant species . The pathogen is highly successful due to its prolific reproductive capacity, remarkable survival mechanisms, and multifaceted infection strategies. This combination of characteristics contributes to its global distribution and has listed it amongst the top 10 most economically devastating phytopathogens .
The life cycle of B. cinerea involves both asexual (B. cinerea) and sexual (Botryotinia fuckeliana) stages. Initially, taxonomic ambiguity existed between the Botryotinia spp. and Botrytis spp.; however, they were demonstrated to represent the different stages of a single fungal species . During the asexual cycle, a mature conidiophore releases conidial spores, which are a major means of disease transmission and primary inoculum. Additionally, B. cinerea forms melanized resting structures known as sclerotia during winter or under unfavourable conditions. Sclerotia, diseased stubbles, and infected seeds can serve as secondary sources of inoculum . Under favourable environmental conditions, the microconidia fertilize sclerotia and form apothecia that undergo meiosis and produce sexual ascospores . However, the sexual cycle is rarely identified in natural environments. Furthermore, the development of sclerotia, conidia, and other developmental stages, as well as the ability to infect hosts, is heavily influenced by factors such as light intensity, light quality, pH, and biological clock. For instance, during continuous darkness and at lower temperatures, the fungus redirects its resources toward sclerotia production, while in light most genotypes tend to produce conidia .


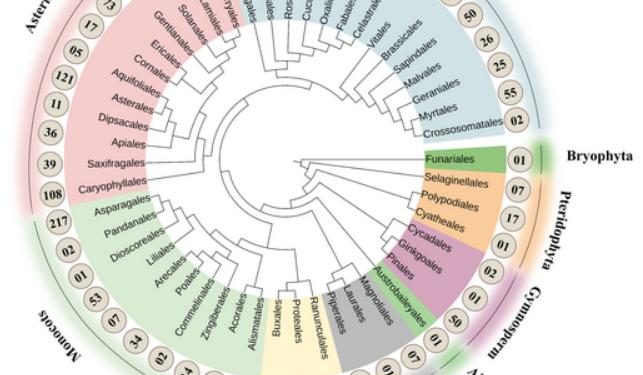
To invade and suppress host defences, the fungus uses sophisticated penetration, infection, and colonization strategies. Infection is primarily initiated by conidia that attach to and germinate on the plant surface. B. cinerea can penetrate host tissues through natural openings such as stomata or wounds. Moreover, to penetrate the host tissue directly, germinating conidia form germ tubes that differentiate into appressoria and infection cushions, specialized structures for host penetration . During the early stage of infection, B. cinerea secretes an arsenal of toxins, cell wall-degrading enzymes, and cell death-inducing proteins along with modulating host apoptotic machinery to facilitate local host cell death . This is the crucial stage in B. cinerea–host interaction, as the balance between B. cinerea and the plant’s cell death pathway determines whether the infection will be halted or progress to the next phase. After successfully establishing inside the host, the fungus expands to surrounding cells. This vigorous growth of B. cinerea is followed by fungal sporulation, production of new conidia, and further disease development. These processes enable B. cinerea to cause pre- and postharvest losses by infecting all parts of its hosts, with organ choice dependent on environmental conditions . In total, B. cinerea infects 616 genera.
B. cinerea has become a model for studying necrotrophic and host generalist mechanisms due its significant impact on global agriculture, broad host range, ease of handling, and ability to infect model plants.
Reference: , & (2024) Genetic and molecular landscapes of the generalist phytopathogen Botrytis cinerea. Molecular Plant Pathology, 25, e13404. Available from: https://doi.org/10.1111/mpp.13404